
Dos trabajos sobre el origen del lenguaje: racionalismo y ciencia
Aquí el término racionalismo no significa uso de la razón, sino razonamiento sin ningún anclaje empírico. En este sentido, los científicos no son racionalistas, por rigurosamente racionales que sean sus pensamientos y por rigurosos que sean sus silogismos. Los dos trabajos que voy a comentar son ejemplos típicos, respectivamente, de racionalismo en el sentido expresado y de ciencia, es decir, de razonamiento con apoyos empíricos sólidos.
En https://rdcu.be/dq8Se, el antropólogo Said Boutiche presenta su visión del origen de las primeras unidades lingüísticas. No explica a qué unidades se refiere, pero es de suponer que a las palabras. Su tesis es que hubo unas presiones selectivas que llevaron al origen de las palabras. Resumiéndolo mucho, esas presiones selectivas se basarían en la vida en grupos sociales amplios y en el intercambio de comida. Esto llevaría a una presión en favor de la equidad que favorecería a los individuos más capaces de redistribuir equitativamente y de conseguir que los otros individuos hicieran lo mismo. Lo cual, a su vez, presionaría en favor del cálculo, que se haría más eficaz si este se podía comunicar claramente. Ahí estaría el origen de las palabras. ¿Pruebas? La distribución de carne en tres sociedades de cazadores-recolectores, con una imagen impactante de uno de sus miembros haciendo y colocando raciones para repartirlas, como vemos en aquí:
No parece que esa prueba pruebe nada más que cómo reparten la caza los cazadores-recolectores actuales y su necesaria capacidad de cálculo, que no parece superar la de los niños de cinco o seis años, ni necesitar ningún tipo de lenguaje humano.
El artículo es un tour de forcé puramente racionalista: los razonamientos pueden estar correctamente construidos, pero no tienen ningún anclaje en los hechos. Esto es más propio del mito que de la ciencia. Los mitos son narraciones inventadas y, como el biólogo y filosofo Henri Atlan puso de manifiesto, pueden tener su propia racionalidad, entendida como subordinación a la lógica, pero a una lógica desencarnada. También son fantásticos. En este caso, lo fantástico es que hace nacer el lenguaje prácticamente de la nada, sin ningún antecedente en todo el reino animal. Esto también es mítico, en el sentido de que habla de un lugar de la humanidad ligado a una supuesta superioridad nuestra. En este sentido, hace afirmaciones cada vez más cuestionadas por la investigación empírica, como que los chimpancés no piensan en futuro ni en lo que no tienen delante, cuando eso se ha demostrado hasta con ratas (https://www.science.org/doi/10.1126/science.adh5206, https://journals.plos.org/ plosbiology/ article?id=10.1371/journal.pbio.3002350).
Diametralmente opuesto es un trabajo de Erich D. Jarvis, neurogenetista especializado en el lenguaje (https://www.science.org/doi/10.1126/science.aax0287). Se centra en el aprendizaje vocal y del lenguaje hablado. Parte de la investigación de las últimas décadas, que apunta a una continuidad entre el lenguaje humano y formas comunicativas vocales de otros animales.
En el apartado en que critica las hipótesis discretas cita casos de estudios que apuntan a la posesión por muchas especies de algunas aptitudes características del lenguaje vocal, añadiendo además que rudimentos de esas capacidades se encuentran también en especies de las que se suponía que no las tenían, como los ratones.
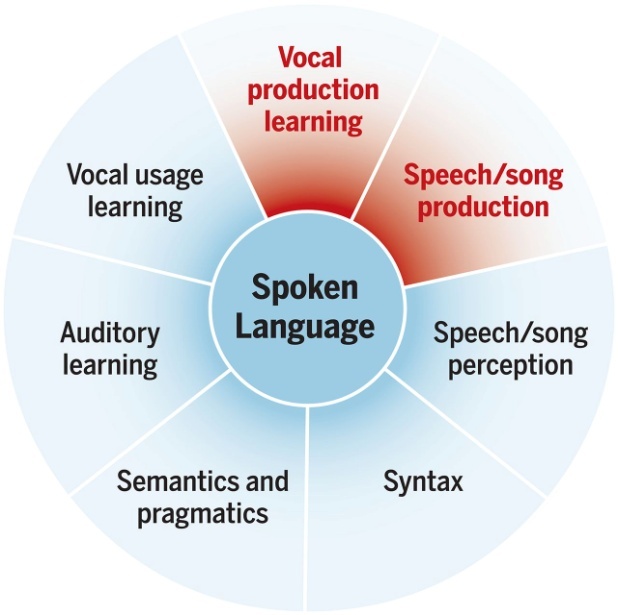
Jarvis propone que el aprendizaje vocal y del habla tiene siete componentes, de los cuales dos están restringidos a muy pocas especies. En un gráfico tomado de Tecumseh Fitch y adaptado para el lenguaje vocal aparecen en rojo el aprendizaje de la producción vocal, restringido a ciertas aves, cetáceos y a los humanos, como lo está la producción del habla/canción. El aprendizaje del uso vocal (en situaciones nuevas) se encuentra, entre otros, en los perros o en los loros. El aprendizaje auditivo lo presentan, por lo menos, todos los mamíferos y todas las aves. Semántica y pragmática son características que presentan los primates (por ejemplo, los monos vervet, con sus cuatro vocalizaciones para avisar de depredadores específicos). La sintaxis se ha encontrado en las aves canoras y en los chimpancés. Y la percepción del habla/canción está bastante extendida entre las mascotas mamíferas y entre las paseriformes y los loros.
La gradación de color en el centro de la figura pretende representar gradientes entre especies en la intensidad de cada componente.
La hipótesis de Jarvis propone una conexión nueva o intensificada entre el córtex motor y las neuronas motoras vocales laríngeas o siríngeas del tronco cerebral en los animales con capacidad de aprendizaje vocal y del habla/canto. Es una de las seis principales hipótesis sobre esta cuestión.
Ni la posición de la laringe, ni la complejidad de su musculatura (la de la siringe) explican el aprendizaje ni la producción de sonidos complejos. Diversas especies de mamíferos sin aprendizaje vocal tienen también la laringe baja y muchas aves no canoras tienen una musculatura siríngea compleja (tal vez en estas especies la falta de aprendizaje vocal esté compensada por la capacidad de producir muchos sonidos innatos). Al parecer, la laringe baja o la siringe compleja serían condiciones necesarias pero no suficientes. Jarvis busca en el encéfalo y en las neuronas.
En la figura de abajo, los esquemas de los cerebros de aves no son hipotéticos. El del cerebro humano sí lo es. Se propone una analogía detallada. Las regiones análogas, coloreadas de la misma manera, incluyen las cortezas motoras laríngeas (LMC) dorsal y ventral responsables de la producción del habla, la LMC premotora y el área de Broca, que intervienen en las funciones superiores del lenguaje hablado. Los núcleos del canto de los pájaros tienen tipos de neuronas que podrían corresponder a las diferentes capas corticales, al cuerpo estriado y a las regiones talámicas de la vía del lenguaje hablado. La entrada a estos circuitos de canto y habla serian vías auditivas, somatosensoriales y de otros tipos que también están presentes en los otros vertebrados, lo que lleva a suponer que son previas a la aparición del canto y del lenguaje. El área de Wernicke habría estado presente en el linaje de los vertebrados desde sus inicios.
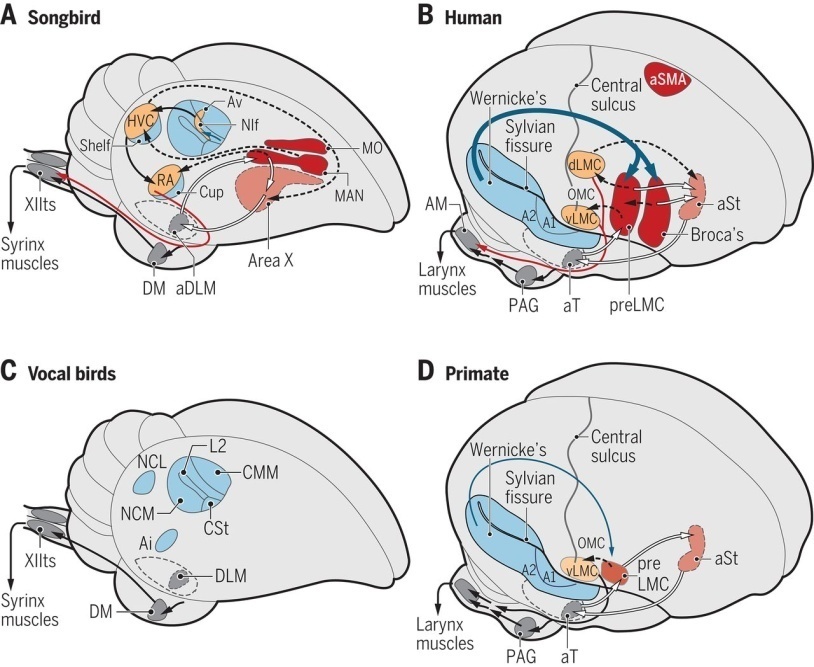
A1: corteza auditiva primaria; A2, corteza auditiva secundaria; aDLM: núcleo medial dorsolateral anterior del tálamo; Ai, arcopallio intermedio; Am, núcleo ambiguo; aSMA: área motora suplementaria anterior; aSt: área del habla del estriado anterior; aT: área del habla del tálamo anterior; Av, avalancha; CMM: mesopalio medial caudal; CSt: estriado caudal; DM: núcleo medial dorsal del mesencéfalo; HVC, un nombre basado en letras; L2, Campo L2; dLMC: corteza motora laríngea dorsal; vLMC: corteza motora laríngea ventral; preLMC: corteza motora laríngea premotora; OMC: corteza motora oral; MAN: núcleo magnocelular del nidopalio; MO: núcleo ovalado de mesopallio; NCM: nidopalio, parte medial caudal; NIf: núcleo interfacial de nidopallium; NLC: nidopalio, caudal lateral; PAG: gris periacueductal; AR, núcleo robusto del arcopallio; XIIts, 12º núcleo motor vocal, parte traqueosirínea. ADAPTADO POR KELLIE HOLOSKI/SCIENCE
La figura siguiente representa la analogía a nivel funcioonal.
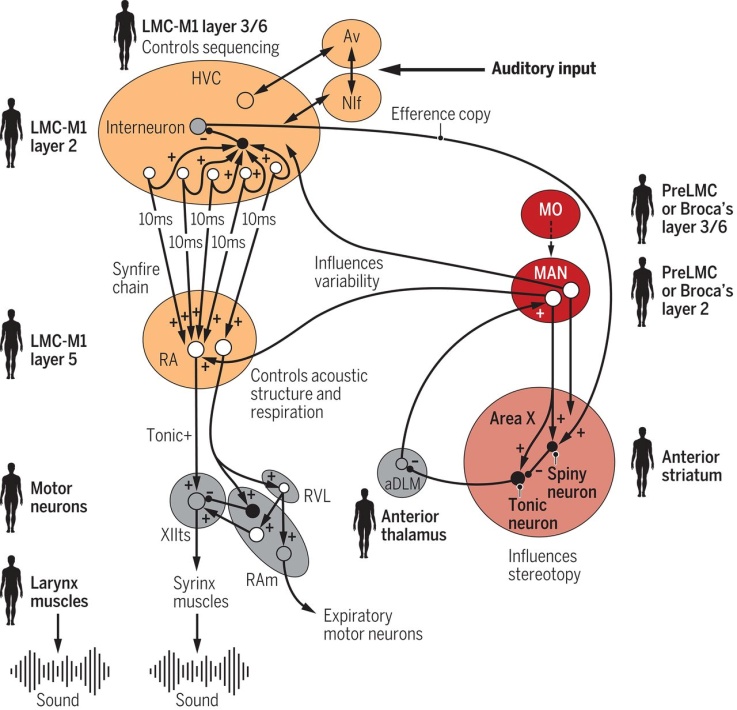
Cada óvalo es un núcleo cantor/vocal o respiratorio de un pájaro cantor. Círculos blancos, neuronas excitadoras; círculos negros, neuronas inhibidoras; signo más, liberación del transmisor excitador; signo menos, liberación del transmisor inhibidor. Las neuronas de proyección RA de HVC se activan en cadenas de disparo sináptico de 10 ms, donde RA traduce esas secuencias para controlar las neuronas premotoras respiratorias y motoras vocales para la producción de cada 10 ms de sonido a través de la siringe (33 ) . MAN y el Área X inyectan variabilidad y estereotopía, respectivamente, tanto en HVC como en RA ( 4 , 35 ). Las regiones del cerebro humano y los tipos de células neuronales previstos que corresponden a los circuitos de aprendizaje vocal en los pájaros cantores están marcados con un contorno humano. Las abreviaturas y el código de colores son los mismos que en la figura anterior . RAm, retroambiguus; RVL, núcleo rostral de la médula ventral-lateral. ADAPTADO POR KELLIE HOLOSKI/ SCIENCE
No es probable que estos circuitos vocales surgieran de novo, sino de duplicaciones de los circuitos motores con posterior especialización. De esa forma se supone que surgieron, por ejemplo, los apéndices bucales de los insectos en los segmentos anteriores de los artrópodos ancestrales.
La hipótesis de Jarvis, de probarse, dejaría como una conjetura muy afortunada la que Calvin y Bickerton enuncian en Lingua ex Machina. Según estos lingüistas, la similitud entre los árboles que describen la formación de oraciones en la gramática generativa de Chomsky y los que describen la preparación del lanzamiento de un proyectil con el brazo, hace pensar que los circuitos lingüísticos descienden de los circuitos motores.
Esta hipótesis, de confirmarse, nos dice mucho más sobre el origen del lenguaje que las decenas de hipótesis, por llamarlas de alguna manera, que tanto gustan a ciertos antropólogos y lingüistas. Y se confirme o no, es pura ciencia, lo que no son hipótesis como la expresada en el artículo que he comentado en primer lugar. Otra diferencia fundamental entre lo que hace Jarvis y lo que hacen esos otros estudiosos es que la hipótesis de Jarvis estimula la investigación, mientras que con esas otras no se puede hacer nada.
Hay que decir dos cosas para acabar. Una, que especies estrechamente emparentadas presentan diferencias de todo o nada en cuanto al aprendizaje vocal, lo que hace pensar que esta característica se ha originado independientemente, es decir, que se ha dado convergencia evolutiva. Y dos, que la hipótesis no hace alusión a presiones selectivas; ya hace bastante, en un tema tan complejo, con establecer posibles vías de evolución.
Abril de 2024