
Los dos principios necesarios de la recapitulación -la adición terminal y la condensación- tenían más fácil explicación, y además una común para los dos, en un marco lamarckiano que en uno darwiniano. Pero su aceptación generalizada como un hecho establecido hizo que los primeros darwinistas se esforzaran en explicarlos en su propio marco teórico.
Las dificultades eran grandes. ¿Cómo explicar mediante la selección natural que la adición terminal en la ontogenia fuera mucho más frecuente que la interpolación (recuérdese que la teoría darwinista considera el proceso evolutivo como un proceso oportunista, de adaptación a entornos locales y muchas veces los entornos de una larva o de un juvenil son muy distintos de los del adulto)? ¿Cómo explicar mediante la selección natural el principio de condensación? Uno podría buscar ventajas adaptativas a la condensación, pero esos argumentos eran débiles y poco convincentes. En vez de esta estrategia, se podría seguir la de aducir que las estructuras se añadían por selección natural, pero las fases de desarrollo eran comprimidas por otro proceso. Lo cual parecía requerir un principio o ley desconocidos de desarrollo, además de reducir el campo de acción de la selección natural.
Müller y otros siguieron la primera estrategia. No había problema en considerar la selección natural como el motor de la adición terminal, siempre y cuando larvas, juveniles y adultos compartieran hábitats y funciones. En cuanto a la condensación, una amplia variación intraespecífica en los ritmos de desarrollo (y planteaba específicamente el caso de la erupción de los dientes en los niños de una misma familia) ofrecía materia prima suficiente a la selección para producir la condensación si un ritmo de desarrollo más rápido era ventajoso. Y consideraba que, en general, la dirección ventajosa era la de condensación, fundamentalmente porque un desarrollo más rápido permitiría acceder antes a un tamaño mayor, con sus efectos de protección y tasas metabólicas más eficientes, y a una madurez sexual más temprana, con la consiguiente posibilidad de dejar más descendencia.
En su favor, aducía Müller que formas relacionadas en diferentes ambientes muestran tasas de condensación muy diferentes relacionadas con sus hábitats. Si la condensación se debiera a una ley de herencia, lo que habría serían tasas de condensación parecidas, reflejo de la similitud genética. También consideraba que cuando las larvas y los adultos vivieran en los mismos ambientes y realizaran las mismas funciones que los adultos, la condensación sería ventajosa. Si vivieran en ambientes distintos, la condensación se frenaría y se interpolarían adaptaciones específicas para los ambientes larvarios y juveniles, como es el caso en las larvas planctónicas de organismos marinos sedentarios, para las cuales sería una ventaja, no sólo la posesión de adaptaciones para la movilidad, sino también que esa fase móvil fuera larga, para tener más oportunidades de encontrar el lugar apropiado para fijarse. Finalmente, afirmaba que la recapitulación predomina porque las ventajas de la condensación son generales.
El más insigne darwinista de finales del XIX, Weismann, que asestó el golpe mortal a la teoría de la herencia de los caracteres adquiridos, no estaba de acuerdo con Müller. Se basaba en su estudio sobre el desarrollo de las pautas de color en las orugas de las Sphingidae, mediante el cual llegó a tres conclusiones:
El desarrollo empieza en un estado de simplicidad y avanza gradualmente hacia la complejidad.
Los caracteres nuevos aparecen en el último estadio de la ontogenia.
Esos caracteres son entonces empujados gradualmente hacia atrás, a estadios ontogenéticos más tempranos, desplazando así los caracteres más antiguos hasta su eventual desaparición completa.
Weismann distinguía tres categorías en las pautas de color de sus orugas: bandas longitudinales, en orugas que viven sobre plantas entre hierba o en la hierba misma; bandas transversales, en orugas que se mueven en hojas de árboles y arbustos; y manchas, con varios significados adaptativos: coloración aposemática (advertencia de incomestibilidad), imitación de ojos (para aterrorizar o sorprender a los predadores) o imitación de bayas.
Dos de estas categorías no tienen valor adaptativo para las orugas muy jóvenes y, por lo tanto de tamaño pequeño, lo que significaría que la explicación del principio de condensación no podía hacerse mediante la selección natural.
La ontogenia general de la coloración de sus orugas apoyaba la primera conclusión: la oruga recién salida del huevo tiene una coloración uniforme. Sus conclusiones segunda y tercera se apoyaban en estudios específicos, la sinopsis de uno de los cuales voy a hacer a continuación.
Se trata del estudio del desarrollo de la pauta de color de las orugas de Chaerocampa elpenor. Weismann observó el siguiente proceso:
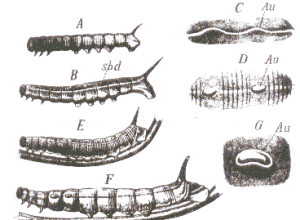
Estrecha banda longitudinal blanca que llamó banda subdorsal (B)
La banda se curva hacia arriba el los segmentos cuarto y quinto (C)
Aparece una línea negra bajo el margen inferior de esas curvas (D)
La línea negra avanza hacia el margen superior.
Finalmente, esa línea se cierra sobre un segmento de la banda blanca, produciendo un "ojo" (G)
Eventualmente, la banda subdorsal desaparece de la mayor parte del resto del cuerpo (F)
La banda subdorsal, en sí misma, podía no tener significado adaptativo, según Weismann, ya que esta oruga no vive entre la hierba sino en grandes hojas de enredadera, pudiendo ser el vestigio de una oruga ancestral que vivía sobre la hierba. Cuando las manchas oculares se desarrollaron como adiciones terminales en la filogenia, la banda longitudinal fue empujada hacia el estadio juvenil en que aparece ahora en la ontogenia.
De ello concluía que no es la selección natural la que causa la condensación, sino, en sus propias palabras, una "ley innata de crecimiento que sigue todo organismo", una especie de ley de herencia aún por descubrir. Veremos en un artículo posterior la paradoja de que el descubrimiento de la ley de herencia fue el golpe de muerte al recapitulacionismo. Pero antes veremos las refutaciones experimentales de la ley biogenética de Haeckel y el papel que jugaron en su derogación, que no fue el que se nos suele dar a entender en las historias de la ciencia al estilo liberal inglés, con su narración del avance constante hacia la verdad a través de los hechos desnudos de la naturaleza.
© Julio Loras Zaera![]()

Julio Loras Zaera
correo e.:
Política de privacidad















![[Validar RSS]](/comun/img/valid-rss.gif "Validar RSS")

![[Valid Atom 1.0]](https://feed1.w3.org/images/valid-rss-rogers.png "Validar Atom 1.0 feed")
